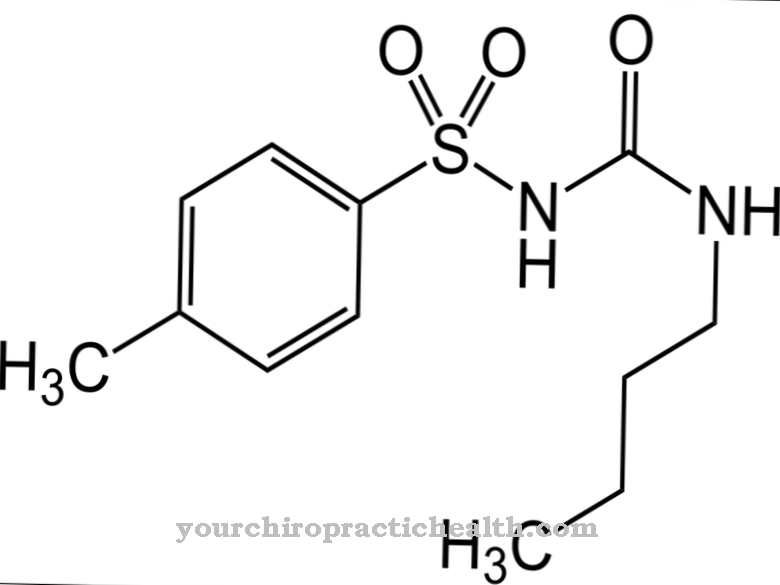
А Декарбоксилювання як правило, являє собою відщеплення діоксиду вуглецю від органічної кислоти. Що стосується карбонових кислот, то відщеплення відбувається дуже добре за допомогою нагрівання та ферментативних реакцій. Окислювальне декарбоксилювання відіграє особливо важливу роль, що призводить до ацетил-КоА в організмі при руйнуванні пірувату та до сукциніл-КоА при руйнуванні α-кетоглутарата.
Що таке декарбоксилювання?

Декарбоксилювання відіграє важливу роль у метаболізмі. Термін декарбоксилювання описує відщеплення вуглекислого газу від органічних молекул. У молекулі вже існує так звана карбоксильна група, яку можна розщепити дією тепла або ферментативних реакцій.
Карбоксильна група містить атом вуглецю, який з’єднаний з атомом кисню подвійною зв'язком, а з гідроксильною групою - єдиним зв’язком.Карбоксильна група замінюється атомом водню гідроксильної групи після відщеплення вуглекислого газу. Наприклад, карбонові кислоти перетворюються на вуглеводні.
Коли розкладаються вуглеводи, жири та білки, загальний баланс катаболічного обміну створює вуглекислий газ, воду та енергію. Вивільнена енергія тимчасово зберігається у формі АТФ і повторно використовується для біологічної роботи, вироблення тепла або для нарощування організмом власних речовин. Декарбоксиляції пірувату та α-кетоглутарату мають величезне значення в контексті метаболізму.
Функція та завдання
Декарбоксилювання відбувається постійно в організмі людини. Важливим субстратом є піруват, який декарбоксилюється за допомогою тіамінпірофосфату (ТЕС). При цьому утворюється гідроксіетил ТЕС (гідроксиетилтіамін пірофосфат) і вуглекислий газ. Ферментом, що відповідає за цю реакцію, є компонент піруватдегідрогенази (Е1).
Тіамін пірофосфат - похідне вітаміну В1. Отриманий в результаті гідроксиетил-ТЕС-комплекс реагує з амідом ліпоєвої кислоти з утворенням ацетилдигідроліпонаміду. Тіамін пірофосфат (ТЕС) утворюється знову. Компонент піруватдегідрогенази також відповідає за цю реакцію.
На наступному етапі ацетилдигідроліпонамід вступає в реакцію з коензимом А з утворенням ацетилу CoA. Фермент дигідроліпоїл трансацетилаза (Е2) відповідає за цю реакцію. Ацетил-КоА являє собою так звану активовану оцтову кислоту. Ця сполука впадає в цикл лимонної кислоти як субстрат і є важливим метаболітом як для анаболічного, так і для катаболічного обміну. Активована оцтова кислота може бути розбита далі на вуглекислий газ і воду або на важливі біологічні субстрати здійснено.
Метаболіт, який вже походить з циклу лимонної кислоти, - α-кетоглутарат. Α-Кетоглутарат також перетворюється за допомогою подібних реакцій з виведенням вуглекислого газу. Створюється кінцевий продукт сукциніл-КоА. Сукциніл-КоА є проміжним продуктом у багатьох обмінних процесах. Він продовжуватиме впроваджуватися як частина циклу лимонної кислоти. Багато амінокислот входять лише в цикл лимонної кислоти через проміжну стадію сукциніл-КоА. Таким чином амінокислоти валін, метіонін, треонін або ізолейцин інтегруються в загальні обмінні процеси.
Загалом, реакції декарбоксилювання пірувату та α-кетоглутарату розташовані на межі між анаболічними та катаболічними обмінними процесами. Вони мають центральне значення для метаболізму. У той же час утворення діоксиду вуглецю шляхом декарбоксилювання включається в загальний баланс вуглекислого газу.
Важливість окисного декарбоксилювання полягає в тому, що в результаті утворюються метаболіти метаболізму, які можуть служити як для отримання енергії для організму, так і для накопичення організмом власних речовин. Декарбоксилювання також відіграє важливу роль у перетворенні глутамату в γ-аміномасляну кислоту (GABA). Ця реакція, каталізується глутаматдекарбоксилазою, є єдиним способом біосинтезу GABA. ГАМК - головний гальмівний нейромедіатор у центральній нервовій системі. Він також відіграє ключову роль у пригніченні гормону підшлункової залози глюкагону.
Хвороби та недуги
Розлади окислювального декарбоксилювання можуть бути спровоковані дефіцитом вітаміну В1. Як вже було сказано, вітамін B1 або його похідне тіамінпірофосфат (TPP) відіграють визначальну роль в окислювальному декарбоксилюванні. Тому дефіцит вітаміну В1 призводить до порушень енергетичного та будівельного обміну. Це призводить до порушення вуглеводного обміну та нервової системи. Може розвинутися полінейропатія. Крім того, виникають симптоми втоми, дратівливості, депресії, порушення зору, погана концентрація, втрата апетиту і навіть атрофія м’язів. Також спостерігаються розлади пам’яті, часті головні болі та анемія.
Імунна система також ослаблена через порушення виробництва енергії. М’язова слабкість в основному впливає на литкові м’язи. Також виникають серцева недостатність, задишка або набряки. У крайній формі дефіцит вітаміну B1 відомий як бери-бери. Бери-бери має місце особливо в регіонах, де раціон дуже низький, а вітамін B1 Це стосується насамперед груп населення, які спеціалізувались на дієтах із соєвими продуктами та ошпареним рисом.
Ще одне захворювання, яке можна простежити до розладу декарбоксиляції, - це так звана спастична тетраплегічна церебральна параліч типу 1. Для цього захворювання, при якому відбувається інфантильний церебральний параліч, тригером є генетичний дефект. Мутація в гені GAD1 призводить до дефіциту ферменту глутаматдекарбоксилази. Глутаматдекарбоксилаза відповідає за перетворення глутамату в γ-аміномасляну кислоту (GABA) шляхом розщеплення вуглекислого газу. Як згадувалося раніше, ГАМК є основним гальмуючим нейротрансмітером центральної нервової системи. Якщо утворюється занадто мало ГАМК, ураження мозку відбувається на ранній стадії. У випадку інфантильного церебрального паралічу вони призводять до спастичного паралічу, атаксиї та атетозу. Спастичний параліч є наслідком постійно підвищеного м’язового тонусу, що призводить до жорсткої постави. У той же час координація рухів порушена у багатьох уражених, що також відоме як атаксія. Крім того, на тлі атетозу можуть спостерігатися мимовільні подовжуючі та химерні рухи, оскільки між гіпотонією та гіпертонією м’язів відбувається постійна зміна.
.jpg)



.jpg)